Биосинтез белка: кратко и понятно. Биосинтез белка в живой клетке
Во всех живых клетках белки синтезируются рибосомами. Рибосома представляет собой крупную макромолекулу со сложной асимметричной четвертичной структурой, построенной из рибонуклеиновых кислот (рибосомных РНК) и белков. Для того чтобы синтезировать белок, рибосома должна быть снабжена:
1. Программой, задающей порядок чередования аминокислотных остатков в полипептидной цепи белка.
2. Аминокислотным материалом, из которого надлежит строить белок.
3. Энергией.
Сама рибосома обладает каталитической (энзиматической) функцией, ответственной за образование пептидных связей и, соответственно, полимеризацию аминокислотных остатков в полипептидную цепь белка.
Программа, задающая порядок чередования аминокислотных остатков в полипептидной цепи белка, исходит от дезоксирибонуклеиновой кислоты (ДНК), т. е.Из генома клетки.Отдельные участки двутяжевойДНК,называемые генами, являются матрицами длясинтеза на них однотяжевых цепей РНК. Синтезированные цепи РНК комплиментарны одной из цепей ДНК и, таким образом, точно воспроизводят дезоксирибонуклеотидную последовательность другой цепи ДНК в своей рибонуклеотидной последовательности. Процесс такого копирования гена, осуществляемый ферментом РНК-полимеразой, получил название транскрипции. РНК в течение синтеза и после него, особенно в эукариотических клетках, может подвергаться ряду дополнительных изменений, называемых процессингом, в ходе которых из нее могут быть вырезаны определенные куски нуклеотидной последовательности. Получающаяся РНК поступает далее в рибосомы в качестве программы, определяющую аминокислотную последовательность в синтезируемом белке. Она называется информационной или "мессенджер" РНК (мРНК). Таким образом, именно транскрипция генов и образование мРНК обеспечивают поток информации от ДНК к рибосомам.
Исходным материалом, из которого строится белок, являются аминокислоты. Однако свободные аминокислоты не используются рибосомой, Для того чтобы служить субстратом для рибосомы, аминокислота должна быть активирована с участием сопряженного расщепления АТФ и акцептирована (ковалентно присоединена) специальной молекулой РНК, называемой трансфернои или транспортной РНК (тРНК), с помощью фермента аминоацил-тРНК-синтетезы. Получающиеся аминоацил-тРНК поступают в рибосому в качестве субстрата для синтеза белка. Кроме того, энергия химической связи между аминокислотным остатком и тРНК используется для реакции образования пептидной связи в рибосоме. Таким образом, активация аминокислот и образование аминоацил-тРНК обеспечивают поток, как материала, так и энергии для рибосомного синтеза белка.
Эти три потока (информации, материала и энергии) встречаются в рибосоме. Воспринимая их, рибосома осуществляет перевод, или трансляцию, генетической информации с языка нуклеотидной последовательности мРНК на язык аминокислотной последовательности синтезируемой полипептидной цепи белка. Если представить это в молекулярных терминах, то рибосома последовательно сканирует цепь мРНК (движется вдоль нее) и тоже последовательно выбирает из среды аминоацил-тРНК, в результате чего специфичность аминоацильного остатка выбираемой рибосомой аминоацил-тРНК каждый раз детерминируется специфичностью комбинации нуклеотидов считываемого в данный момент рибосомой отрезка мРНК. Таким образом, возникает проблема генетического кода: какие комбинации нуклеотидов детерминируют, т. е. Кодируют каждую из 20 аминокислот, из которых строятся молекулы белков?
Движение рибосомы вдоль цепи мРНК (или, другими словами, Пропускание цепи мРНК сквозь рибосому) задает строгий временной порядок вхождения в рибосому разных аминоацил-тРНК в соответствии с порядком расположения кодирующих нуклеотидных комбинаций вдоль мРНК. Аминоацильный остаток выбранной аминоацил-тРНК каждый раз ковалентно присоединяется рибосомой к растущей полипептидной цепи. Деацилированная тРНК освобождается из рибосомы в раствор. Так последовательно, шаг за шагом, строится полипептидная цепь белка (см. схему 1).
© А.С. Спирин
БИОСИНТЕЗ БЕЛКОВ, МИР РНКИ ПРОИСХОЖДЕНИЕ ЖИЗНИ
А.С. Спирин
Спирин Александр Сергеевич - академик, директор Института белка РАН, член Президиума РАН.
Почти полвека тому назад, в 1953 г., Д. Уотсон и Ф. Крик открыли принцип структурной (молекулярной) организации генного вещества - дезоксирибонуклеиновой кислоты (ДНК) . Структура ДНК дала ключ к механизму точного воспроизведения - редупликации - генного вещества . Так возникла новая наука - молекулярная биология. Была сформулирована так называемая центральная догма молекулярной биологии: ДНК Ю РНК Ю белок. Смысл ее состоит в том, что генетическая информация, записанная в ДНК, реализуется в виде белков, но не непосредственно, а через посредство родственного полимера - рибонуклеиновую кислоту (РНК), и этот путь от нуклеиновых кислот к белкам необратим. Таким образом, ДНК синтезируется на ДНК, обеспечивая собственную редупликацию, то есть воспроизведение исходного генетического материала в поколениях; РНК синтезируется на ДНК, в результате чего происходит переписывание, или транскрипция, генетической информации в форму многочисленных копий РНК; молекулы РНК служат матрицами для синтеза белков - генетическая информация транслируется в форму полипептидных цепей. В специальных случаях РНК может переписываться в форму ДНК ("обратная транскрипция"), а также копироваться в виде РНК (репликация), но белок никогда не может быть матрицей для нуклеиновых кислот (подробнее см. ).
Итак, именно ДНК определяет наследственность организмов, то есть воспроизводящийся в поколениях набор белков и связанных с ними признаков. Биосинтез белка является центральным процессом живой материи, а нуклеиновые кислоты обеспечивают его, с одной стороны, программой, определяющей весь набор и специфику синтезируемых белков, а с другой - механизмом точного воспроизведения этой программы в поколениях. Следовательно, происхождение жизни в ее современной клеточной форме сводится к возникновению механизма наследуемого биосинтеза белков.
БИОСИНТЕЗ БЕЛКОВ
Центральная догма молекулярной биологии постулирует лишь путь передачи генетической информации от нуклеиновых кислот к белкам и, следовательно, к свойствам и признакам живого организма. Изучение механизмов реализации этого пути на протяжении десятилетий, последовавших за формулировкой центральной догмы, вскрыло гораздо более разнообразные функции РНК, чем быть только переносчиком информации от генов (ДНК) к белкам и служить матрицей для синтеза белков.
На рис. 1 представлена общая схема биосинтеза белка в клетке.
РНК-посредник
(messenger RNA, матричная РНК, мРНК), кодирующая белки, о
которой и шла речь выше, - это лишь один из трех главных классов клеточных
РНК. Основную их массу (около 80%) составляет другой класс РНК - рибосомные
РНК,
которые образуют структурный каркас и функциональные центры
универсальных белок-синтезирующих частиц - рибосом. Именно рибосомные РНК
ответственны - как в структурном, так и в функциональном отношении - за
формирование ультрамикроскопических молекулярных машин, называемых рибосомами.
Рибосомы воспринимают генетическую информацию в виде молекул мРНК и, будучи
запрограммированы последними, делают белки в точном соответствии с данной
программой.
Однако, чтобы синтезировать белки, одной только информации или программы недостаточно - нужен еще и материал, из которого их можно делать. Поток материала для синтеза белков идет в рибосомы через посредство третьего класса клеточных РНК - РНК-переносчиков (transfer RNA, транспортные РНК, тРНК). Они ковалентно связывают - акцептируют - аминокислоты, которые служат строительным материалом для беЛков, и в виде аминоацил-тРНК поступают в рибосомы. В рибосомах аминоацил-тРНК взаимодействуют с кодонами - трехнуклеотидными комбинациями - мРНК, в результате чего и происходит декодирование кодонов в процессе трансляции.
РИБОНУКЛЕИНОВЫЕ КИСЛОТЫ
Итак, перед нами набор главных клеточных РНК, определяющих основной процесс современной живой материи - биосинтез белка. Это мРНК, рибосомные РНК и тРНК. РНК синтезируются на ДНК с помощью ферментов - РНК-полимераз, осуществляющих транскрипцию - переписывание определенных участков (линейных отрезков) двутяжевой ДНК в форму однотяжевой РНК. Участки ДНК, кодирующие клеточные белки, переписываются в виде мРНК, тогда как для синтеза многочисленных копий рибосомной РНК и тРНК имеются специальные участки клеточного генома, с которых идет интенсивное переписывание без последующей трансляции в белки.
Химическая структура РНК.
Химически РНК очень похожа на ДНК. Оба вещества - это линейные полимеры
нуклеотидов. Каждый мономер - нуклеотид - представляет собой фосфорилированный
N-гликозид, построенный из остатка пятиуглеродного сахара - пентозы, несущего
фосфатную группу на гидроксильной группе пятого углеродного атома
(сложноэфирная связь) и азотистое основание при первом углеродном атоме
(N-гликозидная связь). Главное химическое различие между ДНК и РНК состоит в
том, что сахарный остаток мономера РНК - это рибоза, а мономера ДНК -
дезоксирибоза, являющаяся производным рибозы, в котором отсутствует
гидроксильная группа при втором углеродном атоме (рис. 2).
 Рис. 2. Химические формулы остатков одного из рибонуклеотидов - уридиловой кислоты (U) и гомологичного ему дезоксирибонуклеотида - тимидиловой кислоты (dT) |
Азотистых оснований и в ДНК, и в РНК четыре вида: два пуриновых - аденин (А) и гуанин (G) -и два пиримидиновых - цитозин (С) и урацил (U) или его метилированное производное тимин (Т). Урацил характерен для мономеров РНК, а тимин - для мономеров ДНК, и это второе различие РНК и ДНК. Мономеры - рибонуклеотиды РНК или дезоксирибонуклеотиды ДНК - образуют полимерную цепь посредством формирования фосфодиэфирных мостиков между сахарными остатками (между пятым и третьим атомами углерода пентозы). Таким образом, полимерная цепь нуклеиновой кислоты - ДНК или РНК - может быть представлена как линейный сахаро-фосфатный остов с азотистыми основаниями в качестве боковых групп. Макромолекулярная структура РНК. Принципиальное макроструктурное различие двух типов нуклеиновых кислот состоит в том, что ДНК - это единая двойная спираль, то есть макромолекула из двух комплементарно связанных полимерных тяжей, спирально закрученных вокруг общей оси (см. [ , ]), а РНК - однотяжевой полимер. В то же время взаимодействия боковых групп - азотистых оснований - друг с другом, а также с фосфатами и гидроксилами сахаро-фосфатного остова приводят к тому, что однотяжевой полимер РНК сворачивается на себя и скручивается в компактную структуру , подобно сворачиванию полипептидной цепи белка в компактную глобулу. Таким способом уникальные нуклеотидные последовательности РНК могут формировать уникальные пространственные структуры. |
Впервые специфическая пространственная структура РНК была продемонстрирована при расшифровке атомной структуры одной из тРНК в 1974 г. [ , ] (рис. 3). Сворачивание полимерной цепи тРНК, состоящей из 76 нуклеотидных мономеров, приводит к формированию очень компактного глобулярного ядра, из которого под прямым углом торчат два выступа. Они представляют собой короткие двойные спирали по типу ДНК, но организованные за счет взаимодействия участков одной и той же цепи РНК. Один из выступов является акцептором аминокислоты и участвует в синтезе полипептидной цепи белка на рибосоме, а другой предназначен для комплементарного взаимодействия с кодирующим триплетом (кодоном) мРНК в той же рибосоме. Только такая структура способна специфически взаимодействовать с белком-ферментом, навешивающим аминокислоту на тРНК, и с рибосомой в процессе трансляции, то есть специфически "узнаваться" ими.

Рис. 3. Атомная (слева) и скелетная (справа) модели фенилаланиновой тРНК дрожжей
Изучение изолированных рибосомных РНК дало следующий разительный пример формирования компактных специфических структур из еще более длинных линейных полимеров этого типа. Рибосома состоит из двух неравных частей - большой и малой рибосомных субчастиц (субъединиц). Каждая субчастица построена из одной высокополимерной РНК и целого ряда разнообразных рибосомных белков. Длина цепей рибосомных РНК весьма значительна: так, РНК малой субчастицы бактериальной рибосомы содержит более 1500 нуклеотидов, а РНК большой субчастицы - около 3000 нуклеотидов. У млекопитающих, включая человека, эти РНК еще больше - около 1900 нуклеотидов и более 5000 нуклеотидов в малой и большой субчастицах соответственно.
Было показано, что изолированные рибосомные РНК, отделенные от их белковых
партнеров и полученные в чистом виде, сами способны спонтанно сворачиваться в
компактные структуры, по своим размерам и форме похожие на рибосомные
субчастицы ].
Форма большой и малой субчастиц разная, и соответственно различается форма
большой и малой рибосомных РНК (рис. 4). Таким образом, линейные цепи
рибосомной РНК самоорганизуются в специфические пространственные структуры,
определяющие размеры, форму и, по-видимому, внутреннее устройство рибосомных
субчастиц, а следовательно, и всей рибосомы.
Минорные РНК. По мере изучения компонентов живой клетки и отдельных фракций тотальной клеточной РНК выяснялось, что тремя главными видами РНК дело не ограничивается. Оказалось, что в природе существует множество других видов РНК. Это, в первую очередь, так называемые "малые РНК", которые содержат до 300 нуклеотидов, часто с неизвестными функциями. Как правило, они ассоциированы с одним или несколькими белками и представлены в клетке в виде рибонуклеопротеидов - "малых РНП" .
Малые РНК присутствуют во всех отделах клетки, включая цитоплазму, ядро, ядрышко, ми-тохондрии. Большая часть тех малых РНП, функции которых известны, участвует в механизмах посттранскрипционной обработки главных видов РНК (RNA processing) - превращении предшественников мРНК в зрелые мРНК (сплайсинг), редактировании мРНК, биогенезе тРНК, созревании рибосомных РНК. Один из наиболее богато представленных в клетках видов малых РНП (SRP) играет ключевую роль в транспорте синтезируемых белков через клеточную мембрану. Известны виды малых РНК, выполняющих регуляторные функции в трансляции. Специальная малая РНК входит в состав важнейшего фермента, ответственного за поддержание редупликации ДНК в поколениях клеток - теломеразы. Следует сказать, что их молекулярные размеры сопоставимы с размерами клеточных глобулярных белков. Таким образом, постепенно становится ясно, что функционирование живой клетки определяется не только многообразием синтезируемых в ней белков, но и присутствием богатого набора разнообразных РНК, из которых малые РНК в значительной мере имитируют компактность и размеры белков.
Рибозимы. Вся активная жизнь построена на обмене веществ - метаболизме, и все биохимические реакции метаболизма происходят с надлежащими для обеспечения жизни скоростями только благодаря высокоэффективным специфическим катализаторам, созданным эволюцией. На протяжении многих десятилетий биохимики были уверены, что биологический катализ всегда и всюду осуществляется белками, называемыми ферментами , или энзимами. И вот в 1982-1983 гг. было показано, что в природе имеются виды РНК, которые, подобно белкам, обладают высокоспецифической каталитической активностью [ , ]. Такие РНК-катализаторы были названы рибозимами. Представлению об исключительности белков в катализе биохимических реакций пришел конец.
В настоящее время рибосому тоже принято рассматривать как рибозим. Действительно, все имеющиеся экспериментальные данные свидетельствуют о том, что синтез полипептидной цепи белка в рибосоме катализируется рибосомной РНК, а не рибосомными белками. Идентифицирован каталитический участок большой рибосомной РНК, ответственный за катализ реакции транспептидации, посредством которой осуществляется наращивание полипептидной цепи белка в процессе трансляции.
Что касается репликации вирусных ДНК, то ее механизм мало чем отличается от редупликации генетического материала - ДНК - самой клетки. В случае же вирусных РНК реализуются процессы, которые подавлены или вовсе отсутствуют в нормальных клетках, где вся РНК синтезируется только на ДНК как на матрице. При инфекции РНК-содержащими вирусами ситуация может быть двоякой. В одних случаях на вирусной РНК как на матрице синтезируется ДНК ("обратная транскрипция"), а уж на этой ДНК транскрибируются многочисленные копии вирусной РНК. В других, наиболее интересных для нас случаях на вирусной РНК синтезируется комплементарная цепь РНК, которая и служит матрицей для синтеза - репликации - новых копий вирусной РНК. Таким образом при инфекции РНК-содержащими вирусами реализуется принципиальная способность РНК детерминировать воспроизведение своей собственной структуры, как это имеет место у ДНК.
Мультифункциональность РНК. Суммирование и обзор знаний о функциях РНК позволяют говорить о необыкновенной многофункциональности этого полимера в живой природе. Можно дать следующий список основных известных функций РНК.
Генетическая репликативная функция: структурная возможность копирования (репликации) линейных последовательностей нуклеотидов через комплементарные последовательности. Функция реализуется при вирусных инфекциях и аналогична главной функции ДНК в жизнедеятельности клеточных организмов - редупликации генетического материала.В целом РНК предстает перед нами столь удивительным полимером, что, казалось бы, ни времени эволюции Вселенной, ни интеллекта Творца не должно было бы хватить на ее изобретение. Как можно было видеть, РНК способна выполнять функции обоих принципиально важных для жизни полимеров - ДНК и белков. Неудивительно, что перед наукой и встал вопрос: а не могло ли возникновение и самодостаточное существование мира РНК предшествовать появлению жизни в ее современной ДНК-белковой форме?Кодирующая функция: программирование белкового синтеза линейными последовательностями нуклеотидов. Это та же функция, что и у ДНК. И в ДНК, и в РНК одни и те же триплеты нуклеотидов кодируют 20 аминокислот белков, и последовательность триплетов в цепи нуклеиновой кислоты есть программа для последовательной расстановки 20 видов аминокислот в полипептидной цепи белка.
Структурообразующая функция: формирование уникальных трехмерных структур. Компактно свернутые молекулы малых РНК принципиально подобны трехмерным структурам глобулярных белков, а более длинные молекулы РНК могут образовывать и более крупные биологические частицы или их ядра.
Функция узнавания: высокоспецифические пространственные взаимодействия с другими макромолекулами (в том числе белками и другими РНК) и с малыми лигандами. Эта функция, пожалуй, главная у белков. Она основана на способности полимера сворачиваться уникальным образом и формировать специфические трехмерные структуры. Функция узнавания является базой специфического катализа.
Каталитическая функция: специфический катализ химических реакций рибозимами. Данная функция аналогична энзиматической функции белков-ферментов.
ПРОИСХОЖДЕНИЕ ЖИЗНИ
Белково-коацерватная теория Опарина. Пожалуй, первая научная, хорошо продуманная теория происхождения жизни абиогенным путем была предложена биохимиком А.И. Опариным еще в 20-х годах прошлого века [ , ]. Теория базировалась на представлении, что все начиналось с белков, и на возможности в определенных условиях спонтанного химического синтеза мономеров белков - аминокислот - и белковоподобных полимеров (полипептидов) абиогенным путем. Публикация теории стимулировала многочисленные эксперименты в ряде лабораторий мира, показавшие реальность такого синтеза в искусственных условиях. Теория быстро стала общепринятой и необыкновенно популярной.
Основным ее постулатом было то, что спонтанно возникавшие в первичном "бульоне" белковоподобные соединения объединялись" в коацерватные капли - обособленные коллоидные системы (золи), плавающие в более разбавленном водном растворе. Это давало главную предпосылку возникновения организмов - обособление некой биохимической системы от окружающей среды, ее компартментализацию. Так как некоторые белковоподобные соединения коацерватных капель могли обладать каталитической активностью, то появлялась возможность прохождения биохимических реакций синтеза внутри капель - возникало подобие ассимиляции, а значит, роста коацервата с последующим его распадом на части - размножением. Ассимилирующий, растущий и размножающийся делением коацерват рассматривался как прообраз живой клетки (рис. 5).
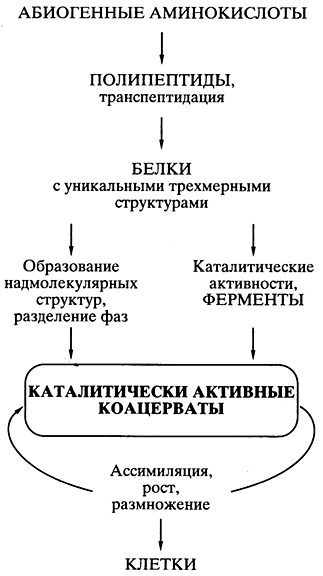
Рис. 5.
Схематическое представление пути
происхождения жизни
согласно белково-коацерватной теории А.И.
Опарина
Все было хорошо продумано и научно обосновано в теории, кроме одной проблемы, на которую долго закрывали глаза почти все специалисты в области происхождения жизни. Если спонтанно, путем случайных безматричных синтезов в коацервате возникали единичные удачные конструкции белковых молекул (например, эффективные катализаторы, обеспечивающие преимущество данному коацервату в росте и размножении), то как они могли копироваться для распространения внутри коацервата, а тем более для передачи коацерватам-потомкам? Теория оказалась неспособной предложить решение проблемы точного воспроизведения - внутри коацервата и в поколениях - единичных, случайно появившихся эффективных белковых структур.
Мир РНК как предшественник современной жизни. Накопление знаний о генетическом коде, нуклеиновых кислотах и биосинтезе белков привело к утверждению принципиально новой идеи о ТОМ, что все начиналось вовсе не с белков, а с РНК [ - ]. Нуклеиновые кислоты являются единственным типом биологических полимеров, макромолекулярная структура которых, благодаря принципу комплементарности при синтезе новых цепей (подробнее см. ), обеспечивает возможность копирования собственной линейной последовательности мономерных звеньев, другими словами, возможность воспроизведения (репликации) полимера, его микроструктуры. Поэтому только нуклеиновые кислоты, но не белки, могут быть генетическим материалом, то есть воспроизводимыми молекулами, повторяющими свою специфическую микроструктуру в поколениях.
По ряду соображений именно РНК, а не ДНК, могла представлять собой первичный генетический материал.
Во-первых, и в химическом синтезе, и в биохимических реакциях рибонуклеотиды предшествуют дезоксирибонуклеотидам; дезоксирибонуклеотиды - продукты модификации рибонуклеотидов (см. рис. 2).
Во-вторых, в самых древних, универсальных процессах жизненного метаболизма широко представлены именно рибонуклеотиды, а не дезоксирибонуклеотиды, включая основные энергетические носители типа рибонуклеозид-полифосфатов (АТФ и т.п.).
В-третьих, репликация РНК может происходить без какого бы то ни было участия ДНК, а механизм редупликации ДНК даже в современном живом мире требует обязательного участия РНК-затравки в инициации синтеза цепи ДНК.
В-четвертых, обладая всеми теми же матричными и генетическими функциями, что и ДНК, РНК способна также к выполнению ряда функций, присущих белкам, включая катализ химических реакций. Таким образом, имеются все основания рассматривать ДНК как более позднее эволюционное приобретение - как модификацию РНК, специализированную для выполнения функции воспроизведения и хранения уникальных копий генов в составе клеточного генома без непосредственного участия в биосинтезе белков.
После того как были открыты каталитически активные РНК, идея первичности
РНК в происхождении жизни получила сильнейший толчок к развитию, и была
сформулирована концепция самодостаточного мира РНК,
предшествовавшего
современной жизни [ ,
].
Возможная схема возникновения мира РНК представлена на рис. 6.
Абиогенный синтез рибонуклеотидов и их ковалентное объединение в олигомеры и полимеры типа РНК могли происходить приблизительно в тех же условиях и в той же химической обстановке, что постулировались для образования аминокислот и полипептидов. Недавно А.Б. Четверин с сотрудниками (Институт белка РАН) экспериментально показали, что по крайней мере некоторые полирибонуклеотиды (РНК) в обычной водной среде способны к спонтанной рекомбинации, то есть обмену отрезками цепи, путем транс-эстерификации . Обмен коротких отрезков цепи на длинные, должен приводить к удлинению полирибонуклеотидов (РНК), а сама подобная рекомбинация способствовать структурному многообразию этих молекул. Среди них могли возникать и каталитически активные молекулы РНК.
Даже крайне редкое появление единичных молекул РНК, которые были способны катализировать полимеризацию рибонуклеотидов или соединение (сплайсинг) олигонуклеотидов на комплементарной цепи как на матрице [ , ], означало становление механизма репликации РНК. Репликация самих РНК-катализаторов (рибозимов) должна была повлечь за собой возникновение самореплицирующихся популяций РНК. Продуцируя свои копии, РНК размножались. Неизбежные ошибки в копировании (мутации) и рекомбинации в самореплицирующихся популяциях РНК создавали все большее разнообразие этого мира. Таким образом, предполагаемый древний мир РНК - это "самодостаточный биологический мир, в котором молекулы РНК функционировали и как генетический материал, и как энзимоподобные катализаторы" .
Возникновение биосинтеза белка. Далее на основе мира РНК должно было происходить становление механизмов биосинтеза белка, появление разнообразных белков с наследуемой структурой и свойствами, компартментализация систем биосинтеза белка и белковых наборов, возможно, в форме коацерватов и эволюция последних в клеточные структуры - живые клетки (см. рис. 6).
Проблема перехода от древнего мира РНК к современному белок-синтезирующему миру - наиболее трудная даже для чисто теоретического решения. Возможность абиогенного синтеза по-липептидов и белковоподобных веществ не помогает в решении проблемы, так как не просматривается никакого конкретного пути, как этот синтез мог бы быть сопряжен с РНК и подпасть под генетический контроль. Генетически контролируемый синтез полипептидов и белков должен был развиваться независимо от первичного абиогенного синтеза, своим путем, на базе уже существовавшего мира РНК. В литературе предложено несколько гипотез происхождения современного механизма биосинтеза белка в мире РНК, но, пожалуй, ни одна из них не может рассматриваться как детально продуманная и безупречная с точки зрения физико-химических возможностей. Представлю свою версию процесса эволюции и специализации РНК, ведущего к возникновению аппарата биосинтеза белка (рис. 7), но и она не претендует на законченность.
Предлагаемая гипотетическая схема содержит два существенных момента, кажущихся принципиальными.
Во-первых, постулируется, что абиогенно синтезируемые олигорибонуклеотиды активно рекомбинировали посредством механизма спонтанной неэнзиматической трансэстерификации , приводя к образованию удлиненных цепей РНК и давая начало их многообразию. Именно этим путем в популяции олигонуклеотидов и полинуклеотидов и могли появиться как каталитически активные виды РНК (рибозимы), так и другие виды РНК со специализированными функциями (см. рис. 7). Более того, неэнзиматическая рекомбинация олигонуклеотидов, комплементарно связывающихся с полинуклеотидной матрицей, могла обеспечить сшивание (сплайсинг) фрагментов, комплементарных этой матрице, в единую цепь. Именно таким способом, а не катализируемой полимеризацией мононуклеотидов, могло осуществляться первичные копирование (размножение) РНК. Разумеется, если появлялись рибозимы, обладавшие полимеразной активностью , то эффективность (точность, скорость и продуктивность) копирования на комплементарной. матрице должна была значительно возрастать.
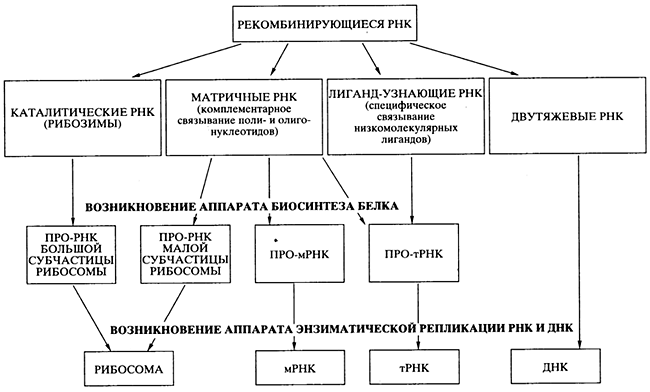
Рис. 7.
Схема эволюции и специализации молекул РНК
в процессе перехода от древнего мира РНК к современному миру
генетически детерминированного биосинтеза белков
Второй принципиальный момент в моей версии состоит в том, что первичный аппарат биосинтеза белка возник на базе нескольких видов специализированных РНК до появления аппарата энзиматической (полимеразной) репликации генетического материала - РНК и ДНК. Этот первичный аппарат включал каталитически активную прорибосомную РНК, обладавшую пептидил-трансферазной активностью; набор про-тРНК, специфически связывающих аминокислоты или короткие пептиды; другую прорибосомную РНК, способную взаимодействовать одновременно с каталитической прорибосомной РНК, про-мРНК и про-тРНК (см. рис. 7). Такая система уже могла синтезировать полипептидные цепи за счет катализируемой ею реакции транспептидации. Среди прочих каталитически активных белков - первичных ферментов (энзимов) - появились и белки, катализирующие полимеризацию нуклеотидов - репликазы, или НК-полимеразы.
Впрочем, возможно, что гипотеза о древнем мире РНК как предшественнике современного живого мира так и не сможет получить достаточного обоснования для преодоления основной трудности - научно правдоподобного описания механизма перехода от РНК и ее репликации к биосинтезу белка. Имеется привлекательная и детально продуманная альтернативная гипотеза А.Д. Альтштейна (Институт биологии гена РАН), в которой постулируется, что репликация генетического материала и его трансляция - синтез белка - возникали и эволюционировали одновременно и сопряженно, начиная с взаимодействия абиогенно синтезирующихся олигонуклеотидов и аминоацил-нуклеотидилатов - смешанных ангидридов аминокислот и нуклеотидов . Но это уже следующая сказка... ("И Шахразаду застигло утро, и она прекратила дозволенные речи" .)
Литература
1. Watson J.D., Crick F.H.C.
Molecular structure of
nucleic acids // Nature. 1953. V. 171. P. 738-740. 2. Watson J.D., Crick F.H.C.
Genetic implications of
the structure of deoxyribose nucleic acid // Nature 1953 V. 171. P. 964-967. 3. Спирин А.С.
Современная биология и биологическая
безопасность // Вестник РАН. 1997. № 7. 4. Spirin A.S.
On macromolecular structure of native
high-polymer ribonucleic acid in solution // Journal of Molecular Biology.
1960. V. 2. P. 436-446. 5. Kirn S.H., Suddath F.L., Quigley GJ. et al.
Three-dimensional tertiary structure of yeast phenylalanine transfer RNA //
Science. 1974. V. 185. P. 435-40. 6. Robertas J.D., Ladner J.E., Finch J.T. et al.
Structure of yeast phenylalanine tRNA at 3 A resolution // Nature. 1974.
V. 250. P. 546-551. 7. Vasiliev V.D., Serdyuk I.N., Gudkov A.T., SPIRin
A.S.
Self-organization of ribosomal RNA // Sturcture, Function and
Genetics of Ribosomes / Eds. Hardesty B. and Kramer G. New York:
Springer-Verlag, 1986. P. 129-142. 8. Baserga SJ., Steitz J.A.
The diverse world of small
ribo-nucleoproteins // The RNA World / Eds. Gesteland R.F. and Atkins J.F. New
York: Cold Spring Harbor Laboratory Press, Cold Spring Harbor, 1993. P.
359-381. 9. Kruger К., Grabowski PJ., Zaug AJ. et al.
Self-splicing RNA: Autoexcision and autocyclization of the ribosomal RNA
intervening sequence of Tetrahymena
// Cell. 1982. V. 31. P. 147-157. 10. Guerrier-Takada С., Gardiner К., Marsh Т. et al.
The RNA moiety of ribonucleases P is the catalytic subunit of the enzyme
// Cell. 1983. V. 35. P. 849-857. 11. Опарин А.И.
Происхождение жизни. М.: Московский
рабочий, 1924. 12. Опарин А.И.
Возникновение жизни на Земле (3-е
изд.). М.: Изд-во АН СССР, 1957. 13. Woese С.
The evolution of the genetic code // The
Genetic Code. New York: Harper & Row, 1967. P. 179-195. 14. Crick F.H.C.
The origin of the genetic code //
Journal of Molecular Biology. 1968. V. 38. P. 367-379. 15. Orgel L.E.
Evolution of the genetic apparatus //
Journal of Molecular Biology. 1968. V. 38. P. 381-393. 16. Gilbert W.
The RNA world // Nature. 1986. V 319
P. 618. 17. Joyce G.F., Orgel L.E.
Prospects for
understanding the origin of the RNA world // The RNA World / Eds. Gesteland
R.F. and Atkins J.F.
New York: Cold Spring Harbor Laboratory Press, Cold
Spring Harbor, 1993 P 1-25. 18. Chetverina H.V., Demidenko A.A., Ugarov V.I.,
Chetverin A.B.
Spontaneous rearrangements in RNA sequences // FEBS
Letters. 1999. V. 450. P. 89-94. 19. Bartel D.P., Szostak J.W.
Isolation of new
ribozymes from a large pool of random sequences // Science. 1993. V. 261. P.
1411-1418. 20. Ekland E.H., Bartel D.P.
RNA-catalysed RNA
polymerization using nucleoside triphosphates // Nature. 1996 V. 382. P.
373-376. 21. Orgel L.E.
The origin of life - a review of facts
and speculations //Trends in Biochemical Sciences. 1998. V. 23. p. 491-495. 22. Альтштейн А.Д.
Происхождение генетической
системы: гипотеза прогенов // Молекулярная биология. 1987. Т. 21. С. 309-322.
Белки играют очень важную роль в жизнедеятельности организмов, выполняют защитные, структурные, гормональные, энергетические функции. Обеспечивают рост мышечной и костной ткани. Белки информируют о строении клетки, о её функциях и биохимических свойствах, входят в состав ценных, полезных организму продуктов питания (яиц, молочных продуктов, рыбы, орехов, бобовых, ржи и пшеницы). Усвояемость такой пищи объясняется биологической ценностью. При равном показателе количества белка легче будет усваиваться тот продукт, чья ценность выше. Дефектные полимеры должны удаляться из организма и заменяться новыми. Этот процесс протекает при синтезе белков в клетках.
Какими бывают белки
Вещества, состоящие только из остатков аминокислот, называются простыми белками (протеинами). В случае необходимости используется их энергетическое свойство, поэтому людям, ведущим здоровый образ жизни, зачастую дополнительно нужен прием протеина. Сложные же белки, протеиды, имеют в своем составе простой белок и небелковую часть. Десять аминокислот в белке являются незаменимыми, это означает, что организм не может синтезировать их самостоятельно, они поступают из пищи, другой же десяток - заменимый, то есть их можно создать из других аминокислот. Так начинается жизненно необходимый для всех организмов процесс.

Основные этапы биосинтеза: откуда берутся белки
Новые молекулы берутся в результате биосинтеза - химической реакции соединения. Существует два основных этапа синтеза белков в клетке. Это транскрипция и трансляция. Транскрипция происходит в ядре. Это считывание с ДНК (дезоксирибонуклеиновой кислоты), которая несет информацию о будущем белке, на РНК (рибонуклеиновую кислоту), которая переносит эту информацию с ДНК в цитоплазму. Происходит это по причине того, что ДНК непосредственно в биосинтезе участия не принимает, она только несет сведения, не имея способности выходить в цитоплазму, где синтезируется белок, и выполняя только функцию носителя генетической информации. Транскрипция же позволяет считать данные с матрицы ДНК на РНК по принципу комплементарности.

Роль РНК и ДНК в процессе
Итак, запускает синтез белков в клетках цепочка ДНК, которая несет информацию о каком-либо конкретном белке и называется геном. Цепочка ДНК в процессе транскрипции расплетается, то есть её спираль начинает распадаться в линейную молекулу. С ДНК информация должна преобразоваться на РНК. Напротив тимина в данном процессе должен становиться аденин. Цитозин же имеет в качестве пары гуанин, точно так же, как ДНК. Напротив аденина РНК становится урацил, потому как в РНК такого нуклеотида, как тимин, не существует, он заменяется просто урациловым нуклеотидом. С гуанином соседствует цитозин. Напротив аденина становится урацил, а в паре с тимином располагается аденин. Эти молекулы РНК, которые становятся напротив, называются информационными РНК (иРНК). Они способны через поры выходить из ядра в цитоплазму и рибосомы, которые, собственно, и выполняют функцию синтеза белков в клетках.
О сложном простыми словами
Теперь же совершается сборка из аминокислотных последовательностей полипептидной цепочки белка. Транскрипцией можно назвать считывание информации о будущем белке с матрицы ДНК на РНК. Это можно определить как первый этап. После того как РНК выходит из ядра, она должна попасть к рибосомам, где происходит второй этап, который называется трансляцией.

Трансляция - это уже переход РНК, то есть перенос информации с нуклеотидов на молекулу белка, когда РНК говорит о том, какая последовательность аминокислот должна быть в веществе. В таком порядке информационная РНК попадает в цитоплазму к рибосомам, которые осуществляют синтез белков в клетке: А (аденин) - Г (гуанин) - У (урацил) - Ц (цитозин) - У (урацил) - А (аденин).
Зачем нужны рибосомы
Для того чтобы произошла трансляция и в результате образовался белок, нужны такие компоненты, как сама информационная РНК, транспортная РНК, а также рибосомы в качестве "фабрики", на которой производится белок. В данном случае функционируют две разновидности РНК: информационная, которая образовалась в ядре с ДНК, и транспортная. Молекула второй кислоты имеет вид клевера. Этот "клевер" присоединяет к себе аминокислоту и несет её к рибосомам. То есть он выполняет транспортировку органических соединений непосредственно к "фабрике" по их образованию.
Как работает рРНК
Также существуют рибосомальные РНК, которые входят в состав самой рибосомы и выполняют синтез белка в клетке. Получается, что рибосомы являются немембранными структурами, они не имеют оболочек, как, например, ядро или эндоплазматическая сеть, а состоят просто из белков и рибосомальных РНК. Что же происходит, когда последовательность из нуклеотидов, то есть информационная РНК, попадает к рибосомам?

Транспортная РНК, которая находится в цитоплазме, подтягивает к себе аминокислоты. Откуда же взялись аминокислоты в клетке? А образуются они вследствие расщепления белков, которые поступают внутрь с пищей. Эти соединения переносятся током крови к клеткам, где происходит продуцирование необходимых для организма белков.
Конечный этап синтеза белков в клетках
Аминокислоты плавают в цитоплазме так же, как и транспортные РНК, и когда происходит непосредственно сборка полипептидной цепи, эти транспортные РНК начинают с ними соединяться. Однако не во всякой последовательности и далеко не любая транспортная РНК может соединиться со всеми видами аминокислот. Существует определенный участок, к которому присоединяется необходимая аминокислота. Второй же участок транспортной РНК называется антикодоном. Этот элемент состоит из трех нуклеотидов, которые комплементарны последовательности нуклеотидов в информационной РНК. Для одной аминокислоты необходимо три нуклеотида. Например, какой-либо условный белок состоит для упрощения из всего лишь двух аминокислот. Очевидно, что в основном белки имеют очень длинную структуру, состоят из многих аминокислот. Цепь А - Г - У называется триплетом, или кодоном, к нему будет присоединяться транспортная РНК в виде клевера, на конце которого будет находиться определенная аминокислота. К следующему триплету Ц - У - А будет присоединяться еще одна тРНК, которая будет содержать совершенно другую аминокислоту, комплементарную данной последовательности. В таком порядке будет происходить дальнейшая сборка полипептидной цепочки.

Биологическое значение синтеза
Между двумя аминокислотами, находящимися на концах "клеверов" каждого триплета, образуется пептидная связь. На этом этапе транспортная РНК уходит в цитоплазму. К триплетам присоединяется затем следующая транспортная РНК с другой аминокислотой, которая образует с предыдущими двумя полипептидную цепь. Этот процесс повторяется до момента, когда набирается необходимая последовательность аминокислот. Таким образом происходит синтез белка в клетке, и образуются ферменты, гормоны, кровяные вещества и т. д. Не во всякой клетке образуется любой белок. Каждая клетка может образовать определенный белок. Например, в эритроцитах будет образовываться гемоглобин, а клетками поджелудочной железы будут синтезироваться гормоны и разнообразные ферменты, расщепляющие пищу, которая попадает в организм.
В мышцах же будет образовываться белок актин и миозин. Как видно, процесс синтеза белка в клетках многоэтапен и сложен, что говорит о его значимости и необходимости для всего живого.
Биосинтез белков (полипептидов) является чрезвычайно сложным и удивительным процессом. Биосинтез белков активно протекает во всех органах и тканях, исключая эритроциты. Многие клетки синтезируют белки на «экспорт» (клетки печени, поджелудочной железы), и в этом случае они содержат очень большое число рибосом. В животной клетке число рибосом достигает 10 5 , диаметр рибосомы равен 20 нм.
Процесс синтеза белка происходит внутри клеток на поверхности рибосом, которые представляют собой комплексы из двух субъединиц с константой седиментации 60S и 40S, функционирующих как единое целое. В рибосоме белок составляет 30-35% и рибосомальная РНК - 65-70%. В рибосоме различают аминоацильный и пептидильный участки. Первый служит для фиксации поступающего на рибосому комплекса активной аминокислоты и тРНК, а второй фиксирует полипептидную цепь, связанную с другой тРНК. Субъединицы рибосом синтезируются в ядрышке ядра на матрице ДНК.
Сущность процесса синтеза белка представляет схема:
Белоксинтезирующая система включает рибосомы, нуклеиновые кислоты, набор из 20 аминокислот, различные ферменты, АТФ, ГТФ, ионы магния, около 200 различных некаталитических белковых факторов.
Молекула белка - длинная цепь аминокислотных остатков, насчитывающая в среднем от 100 до 500 аминокислот. Программа синтеза каждого белка хранится в молекуле дезоксирибонуклеиновой кислоты (ДНК). Молекула ДНК - полимер, мономерами которого служат нуклеотиды. Последовательность азотистых оснований в молекуле ДНК определяет последовательность аминокислот в молекуле белка.
В молекуле ДНК имеются четыре вида азотистых оснований: аденин (А), гуанин (Г), цитозин (Ц) и тимин (Т). Последовательность из трех оснований (триплет) составляет кодон, которому соответствует одна определенная аминокислота.
Нуклеиновые кислоты - ДНК и РНК - обязательные компоненты биосинтеза белков. ДНК отвечает за сохранение генетической информации, тогда как РНК определяет передачу этой информации и реализацию в виде молекул белка. Можно утверждать, что главная функция ДНК - это сохранение генотипа, а РНК - выражение этого генотипа.
В количественном плане в клетке преобладает рибосомаль- ная РНК (рРНК). рРНК имеет спирализованные участки, содержит модифицированные нуклеотиды (например, 2-метил- рибоза). рРНК составляет около 80% от общего количества РНК в клетке. Второй вид РНК в клетке представлен транспортной РНК (тРНК), которая, как и все другие виды РНК, синтезируется в ядре. На ее долю приходится 10-15% общего количества РНК в клетке. Выявлено свыше 60 различных тРНК. Поэтому для транспорта отдельных аминокислот существует несколько разных тРНК. Для каждой аминокислоты в клетке есть по крайней мере одна специфическая тРНК. Молекулы тРНК сравнительно мелкие. В их структуре 75-93 рибонук- леотидов.
Аминокислота присоединяется к свободной 3-ОН-группе концевого мононуклеотида тРНК, представленной всегда аде- ниловой кислотой. тРНК имеет и другой важный участок - антикодон, с помощью которого комплекс аминокислоты и тРНК узнает определенную последовательность из трех нуклеотидов в матричной РНК (кодон). Антикодон и кодон комплементарно соединяются посредством водородных связей.
Если носителем наследственной информации в клетке является ДНК, которая сосредоточена в ядре, но синтез белка происходит в цитоплазме, то, следовательно, должен быть определенный посредник, передающий эту информацию в цитоплазму клетки. Этим посредником оказалась информационная или матричная РНК (мРНК). На долю мРНК приходится 2% общего количества РНК клетки. Молекулы мРНК самые длинные (включают до 5 тыс. нуклеотидов). мРНК также содержит четыре вида азотистых оснований. Из них три (А, Г, Ц) такие же, как в ДНК, а четвертое - урацил.
Информация, закодированная в мРНК, необходима для синтеза молекулы белка, который происходит на рибосомах. Синтез мРНК в ядре клетки очень быстрый, что необходимо для активного биосинтеза белковых молекул. мРНК образуется на одной из нитей ДНК ядра. При этом двухспиральная структура ДНК раскручивается и при участии ДНК-зависимой РНК-по- лимеразы по принципу комплементарности происходит синтез мРНК:

Схема синтеза мРНК
Принцип комплементарности означает, что аденину на спирали ДНК соответствует урацил мРНК, тимину - аденин, а гуанину - цитозин. Следовательно, мРНК считывает информацию с ДНК.
Стадия ДНК -» РНК, таким образом, определяет синтез молекулы мРНК, в которой нуклеотидная последовательность комплементарна определенному участку (гену) ДНК. Этот процесс носит название транскрипции. Затем мРНК поступает на рибосому, объединяясь с ее субъединицами. Одна молекула мРНК фиксируется на множестве рибосом одновременно, образуя так называемые полисомы. Наличие полисом повышает эффективность и скорость использования мРНК.
Синтез полипептидной цепи определенного состава происходит на матрице мРНК. Процесс передачи информации с мРНК на белок получил название трансляции. Стадия «РНК -> белок » представляет процесс синтеза белка, направляемый мРНК. Таким образом, передача информации всегда идет в направлении ДНК -» РНК -» белок.
Процесс трансляции включает следующие этапы:
- 1) активация аминокислот и их фиксация на тРНК;
- 2) инициация синтеза полипептидной цепи;
- 3) элонгация синтезируемой полипептидной цепи;
- 4) терминация полипептидной цепи и ее освобождение;
- 5) посттрансляционная модификация полипептидной цепи.
- 1. Активация аминокислот требует фермента аминоацил- тРНК-синтетазы и затраты энергии в виде АТФ:
Этот же фермент участвует в фиксации предварительно активированной аминокислоты в положение 2 или 3 рибозы последнего нуклеотида тРНК:
В виде данного комплекса аминокислота транспортируется на рибосому, на которой происходит синтез белковой молекулы. Аминоацил-тРНК-синтетаза специфична, она способна узнавать как аминокислоту, так и тРНК. В клетке, таким образом, имеется не менее 20 различных синтетаз, в соответствии с числом а-аминокислот.
2. тРНК, связанная эфирной связью с определенной аминокислотой, поступает на рибосому и взаимодействует с мРНК по типу комплементарности между специфическим триплетом нуклеотидов мРНК, названным кодоном, и ей комплементарным специфическим триплетом нуклеотидов (антикодоном) тРНК, переносящей определенную аминокислоту. Таким образом, каждый кодон мРНК соответствует специфической фиксации одной аминокислоты в пептидной цепи посредством антикодона тРНК. Рибосома передвигается вдоль молекулы мРНК, считывая последовательно все кодоны, устанавливая таким образом порядок расположения всех аминокислот, доставляемых к месту синтеза.
Синтез молекулы белка идет по направлению от свободной аминогруппы к свободной карбоксильной группе аминокислоты. Обычно начальной аминокислотой в синтезе полипептид- ной цепи является метионин, для которой кодоном служит нуклеотидная последовательность АУГ мРНК.
Инициация синтеза полипептида начинается при фиксации двух антикодонов тРНК по соответствующим кодонам мРНК. Процесс требует наличия источника энергии, которым служит ГТФ, а также участия целого ряда белковых факторов инициации и пептидилтрансферазы.
При участии данного фермента скорость образования ковалентных связей достигает 1200 аминокислот/мин/рибосому.

Схема инициации синтеза полипептида
3. После образования дипептида «ненагруженная» тРНК покидает рибосому и способна доставлять новые молекулы аминокислот, а мРНК продвигается относительно рибосомы (полисомы) на три нуклеотида. В результате перемещения (транслокации) свободный кодон занимает положение для узнавания очередной молекулы тРНК. Следовательно, в стадии элонгации происходит последовательное присоединение по одной аминокислоте к полипептидной цепи в строгом соответствии с порядком кодонов молекулы мРНК.
Удлиняющаяся полипептидная цепь с одной молекулой тРНК фиксируется с большой субъединицей рибосомы. Присоединение каждой дополнительной аминокислоты к полипептидной цепи происходит за счет взаимосвязи аминогруппы присоединяющейся аминокислоты в комплексе с тРНК и карбоксильной группы пептида.
4. Терминация, или завершение синтеза полипептидной молекулы, вовлекает определенные кодоны терминации «без смысла» и белковые факторы терминации. Известны три кодона (УАГ, УГА, УАА), которые не кодируют, не связывают какую-либо аминокислоту, так как в клетке не существует антикодонов тРНК, комплементарных к ним. Теоретически лишь один кодон «без смысла», узнаваемый полисомой во время прохождения в направлении 5-3 мРНК, должен остановить синтез молекулы белка.
Наличие терминирующего кодона в любом участке мРНК означает окончание белкового синтеза. В результате полисома распадается, неиспользованная мРНК гидролизуется полинук- леотидфосфорилазой, а субъединицы рибосом готовятся к началу синтеза новой молекулы белка.
мРНК может неоднократно участвовать в процессе биосинтеза белка. Продолжительность функционирования молекулы мРНК неодинакова у различных организмов. Она может колебаться от нескольких минут до нескольких суток.
5. В ДНК закодирована лишь первичная структура белка. Поэтому синтезированные на рибосомах молекулы белков еще не имеют окончательно завершенного состояния. Они представляют первичные полипептиды, которые затем претерпевают многочисленные модификации (ассоциации мономеров с образованием олигомеров, присоединения коферментов, химические превращения), изменяющие структуру белков и, значит, их активность.
Вторичная и третичная структуры не кодированы, они определяются свойствами первичной структуры, а это значит, что та или иная форма белковой молекулы зависит от последовательности аминокислот и возможностей их взаимодействия между собой. Структурные модификации синтезируемых белков имеют место еще на уровне рибосом или после завершения синтеза в результате присоединения различных функциональных групп.
Рассмотренная схема передачи информации в виде
может в отдельных случаях изменяться. Так, у вирусов, не содержащих ДНК, информация заложена в РНК. При проникновении вируса в клетку эта информация передается на ДНК клетки, а последняя уже синтезирует мРНК, на матрице которой синтезируются вирусные белки. Такой процесс носит название обратной транскрипции, и схема передачи информации в этом случае будет следующей:
Пока сохраняется последовательность нуклеотидов ДНК и, следовательно, мРНК, характер вновь синтезируемого белка остается неизменным.
Необходимая генетическая информация для синтеза белка может быть представлена аналогично записи человеческого языка, которая состоит из последовательности букв, формирующих слова и предложения. В генетическом языке, однако, есть только четыре буквы - четыре основания (аденин, гуанин, урацил, цитозин).
Генетический код включает трехбуквенные слова. Четыре основания в данном случае (43) дают 64 варианта (слова), которых более чем достаточно, чтобы кодировать 20 аминокислот. Таким образом, 64 кодона и составляют генетический код (табл. 3).
Анализ генетического кода показывает, что для различных аминокислот имеется различное число кодонов. Например, метионин и триптофан имеют только один кодон, тогда как аргинин, лейцин, серин имеют по шесть кодонов. Наличие нескольких кодонов для одной аминокислоты отражает «вырожден- ность» кода. Следовательно, одна и та же аминокислота может кодироваться несколькими по своему строению нуклеотидными триплетами. В то же время каждому триплету соответствует вполне определенная аминокислота в синтезируемой поли- пептидной цепи.
Т а б л и ц а 3
Генетический код
|
нуклеотид |
Второй нуклеотид |
нуклеотид |
|||
Генетический код универсален и одинаков у видов разного уровня развития (человек, животные, растения, микроорганизмы). Универсальность кода свидетельствует, что все живые организмы в прошлом имели единого предка.
Отдельные аминокислоты (оксипролин, оксилизин), например, не имеют кодона и образуются с помощью химических реакций уже после синтеза полипептидной цепи. Этот процесс получил название посттрансляционной модификации и очень важен для правильного функционирования каждого белка.
Бессмысленные кодоны (УАА, УАГ, УГА) не кодируют аминокислоты, однако реально служат сигналом окончания синтеза белковой молекулы.
Таким образом, мРНК является непосредственным переносчиком генетической информации из ядра на рибосому цитоплазмы. Одна рибосома занимает на мРНК участок длиной около 80 нуклеотидов и способна катализировать примерно 100 пептидных связей в минуту (Северин Е. С. и др., 2011).
Синтезированные белковые молекулы могут подвергаться структурным модификациям еще на уровне рибосом или после завершения синтеза в результате присоединения различных функциональных групп. В цитоплазме мРНК имеет сравнительно короткий период существования. Некоторое количество мРНК синтезируется и запасается в неактивной форме, будучи готовой для быстрого синтеза белка. Поскольку информация мРНК связана с линейной последовательностью нуклеотидов, целостность этой последовательности чрезвычайно важна. Любая потеря или изменение порядка нуклеотидов может видоизменить синтез белка. На сегодня установлен целый ряд ингибиторов репликации ДНК в клетках организма (антибиотики, химические яды, антивирусные препараты). Повреждения в последовательности пуриновых или пиримидиновых оснований в гене получили название мутации.
Замена лишь одного нуклеотида в кодоне (мутация) приводит к смене кодирования одной аминокислоты на другую. Например, мутация, связанная с заменой глутаминовой кислоты на валин в молекуле гемоглобина, приводит к синтезу гемоглобина, вызывающего серповидную анемию. Сегодня известно более 200 мутаций полипептидной цепи молекулы гемоглобина человека. Часто мутагенами являются вещества (нитроза- мины, например), изменяющие структуру азотистых оснований, что приводит к изменению характера комплементарности оснований. Ультрафиолетовое облучение вызывает конденсацию остатков тимина с образованием тиминовых димеров. К счастью, от вредного действия ультрафиолетовых лучей животные защищены слоем озона атмосферы.
Многие антибиотики, используемые в ветеринарной практике, ингибируют бактериальный синтез белка (линкомицин, эритромицин, хлорамфеникол) еще на стадии трансляции. При этом микробная клетка погибает или приостанавливает свое развитие. Такие антибиотики, как тетрациклины, не влияют на рибосомальный синтез в клетках высших животных. Пени- циллины не являются прямыми ингибиторами синтеза белка, однако их эффекты ингибирования бактерий связаны с блокированием синтеза гексапептидов клеточной стенки. Следует отметить, что синтез белка происходит не только на рибосомах, но и в митохондриях. Митохондрии имеют полный и независимый аппарат синтеза белка для своих нужд, хотя не все митохондриальные белки синтезируются в этих органеллах. РНК митохондрий составляют лишь 3% от всего количества РНК клетки. Рибосомы митохондрий меньше по размерам, чем цитоплазматические. Кодон УГА, как терминатор синтеза белка в цитоплазме, используется в митохондриях наряду с кодоном УГГ для кодирования аминокислоты.
Синтезированные на рибосомах белки еще не имеют окончательно завершенного состояния. Они представляют первичные полипептиды, которые затем претерпевают многочисленные модификации (ассоциации мономеров с образованием олигомеров, присоединения коферментов, химические превращения), модифицирующие структуру белка и, значит, его активность.
Одно из наиболее важных и характерных свойств живой клетки. Первичная структура белка, как уже отмечалось, предопределяется генетическим кодом, заложенным в молекуле ДНК, причем различные ее участки кодируют синтез разных белков. Следовательно, одна молекула ДНК хранит информацию о структуре многих белков.
Свойства белка зависят от последовательности расположения аминокислот в полипептидной цепи. В свою очередь чередование аминокислот определяется последовательностью нуклеотидов в ДНК. В иРНК каждой аминокислоте соответствует определенный триплет - группа, состоящая из трех нуклеотидов, называемая кодоном.
Биосинтез белка начинается в ядре со списывания информации о структуре белковой молекулы с ДНК на иРНК по принципу комплементарности. Данный процесс протекает как реакция матричного синтеза и называется транскрипцией (рис. 7.1).
Рис. 7.1. Процесс транскрипции
В результате транскрипции образуется «незрелая» иРНК (пре-иРНК), которая проходит стадию созревания или процессинга .
Процессинг включает в себя:
1) КЭПирование 5"-конца;
2) полиаденилирование 3"-конца (присоединение нескольких десятков адениловых нуклеотидов);
3) сплайсинг (вырезание интронов и сшивание экзонов). В зрелой иРНК выделяют КЭП, транслируемую область (сшитые в одно целое экзоны), нетранслируемые области (НТО) и полиА «хвост». Возможен альтернативный сплайсинг , при котором вместе с интронами вырезаются и экзоны. При этом с одного гена могут образовываться разные белки. Таким образом, утверждение – «Один ген – один полипептид» – неверно (рис. 7.2, 7.3, 7.4)
Рис. 7.2. Сплайсинг
Рис. 7.3. Альтернативный сплайсинг (варианты)
Рис. 7.4. Образование разных молекул белка при вариантах альтернативного сплайсинга
Образующаяся при этом иРНК поступает в цитоплазму, где на нее нанизываются рибосомы. Одновременно в цитоплазме с помощью ферментов активизируется транспортная РНК тРНК.
Молекула тРНК напоминает по структуре лист клевера, на вершине которого находится триплет нуклеотидов, соответствующий по коду определенной аминокислоте (антикодон), а основание («черешок») служит местом присоединения этой аминокислоты. В тРНК различают антикодоновую петлю и акцепторный участок. В антикодоновой петле РНК имеется антикодон, комплементарный кодовому триплету определенной аминокислоты, а акцепторный участок на 3"-конце способен с помощью фермента аминоацил-тРНК-синтетазы присоединять именно эту аминокислоту (с затратой АТФ) к участку ССА (рис. 5)
Транспортная РНК доставляет аминокислоты к рибосомам. По принципу комплементарности антикодон связывается со своим кодоном, причем аминокислота располагается у активного центра рибосомы и с помощью ферментов соединяется с ранее поступившими аминокислотами. Затем тРНК освобождается от аминокислоты, а молекула иРНК продвигается вперед на один триплет, и процесс повторяется.
Рис. 7.5. Строение молекулы Т-РНК
Различают три этапа в биосинтезе белка: инициацию , элонгацию и терминацию .
В малой субъединице рибосомы расположен функциональный центр рибосомы (ФЦР) с двумя участками - пептидильным (Р-участок) и аминоацильным (А-участок). В ФЦР может находиться шесть нуклеотидов иРНК, три - в пептидильном и три - в аминоацильном участках.
Инициация . Синтез белка начинается с того момента, когда к 5"-концу иРНК присоединяется малая субъединица рибосомы, в Р-участок которой заходит метиониновая тРНК.
За счет АТФ происходит передвижение инициаторного комплекса (малая субъединица рибосомы, тРНК с метионином) по НТО до метионинового кодона АУГ. Этот процесс называется сканированием .
Элонгация . Как только в Р-участок сканирующего комплекса попадает кодон АУГ, происходит присоединение большой субъединицы рибосомы. В А-участок ФЦР поступает вторая тРНК, чей антикодон комплементарно спаривается с кодоном иРНК, находящимся в А-участке.
Пептидилтрансферазный центр большой субъединицы катализирует образование пептидной связи между метионином и второй аминокислотой. Отдельного фермента, катализирующего образование пептидных связей, не существует. Энергия для образования пептидной связи поставляется за счет гидролиза ГТФ.
На один цикл расходуется 2 молекулы ГТФ. В А-участок заходит третья тРНК, и образуется пептидная связь между второй и третьей аминокислотами. Синтез полипептида идет от N-конца к С-концу, то есть пептидная связь образуется между карбоксильной группой первой и аминогруппой второй аминокислоты.
Скорость передвижения рибосомы по иРНК - 5–6 триплетов в секунду, на синтез белковой молекулы, состоящей из сотен аминокислотных остатков, клетке требуется несколько минут.
Терминация . Когда в А-участок попадает кодон-терминатор (УАА, УАГ или УГА), с которым связывается особый белковый фактор освобождения, полипептидная цепь отделяется от тРНК и покидает рибосому. Происходит диссоциация, разъединение субъединиц рибосомы.
Рис. 7.6. Процесс трансляции (шаг 1)
Рис. 7.7. Процесс трансляции (шаг 2)
Рис. 7.8. Процесс трансляции (шаг 3)
Рис. 7.9. Процесс трансляции (шаг 4)
Рис. 7.10. Биосинтез белка (общая схема)
Так постепенно наращивается белковая цепочка, в которой аминокислоты располагаются в строгом соответствии с локализацией кодирующих их триплетов в молекуле иРНК. Синтез полипептидных цепей белков по матрице иРНК называется трансляцией (рис. 10).
В клетках растительных и животных организмов белки непрерывно обновляются. Интенсивность синтеза тех или иных специфических белков определяется активностью соответствующих генов, с которых «считывается» иРНК. Следует отметить, что не все гены функционируют одновременно: активность проявляют лишь те, которые кодируют информацию о структуре белков, необходимых для жизнедеятельности организма в данный момент.